赤血球は酸素を運搬する作用を備えており〔Minias 2020〕、それぞれ異なった生態における鳥種では、渡り戦略、高度、代謝率などの様々な生活史戦略に赤血球が適応していることが予想されます〔Minias et al.2013,Yap et al.2019,Minias 2020〕。外温性の脊椎動物 (魚類、爬虫類、両生類) は内温性の哺乳類や鳥類よりも細胞が大きく〔Glomski and Pica 2006〕、進化にしたがって赤血球のサイズを小さくして数を増やした傾向があります〔Hawkey et al.1991〕。両生類や鳥類(Kozłowski et al.2010)および爬虫類の有鱗目は、分類群内での赤血球サイズの差は体重に比例しますが、赤血球サイズは同種類内の個体間でもばらつきが見られます(爬虫類:Starostová et al.2013、魚類:Lahnsteiner 2020)〔Hawkey et al.1991〕。これは、性、年齢、栄養状態、 妊娠、運動などの影響を受ける可能性があります〔Pis 2008〕。さらに、赤血球サイズは季節的な高度勾配の上下移動〔Janiga et al. 2017〕、温度〔Ruiz et al. 2004〕、環境汚染物質への反応〔Janiga et al.2019〕、および換羽〔Haas et al.2020〕や交尾などのエネルギー需要の年間変動と関連した可塑性変化を示すことがあります。
鳥の赤血球サイズの変化
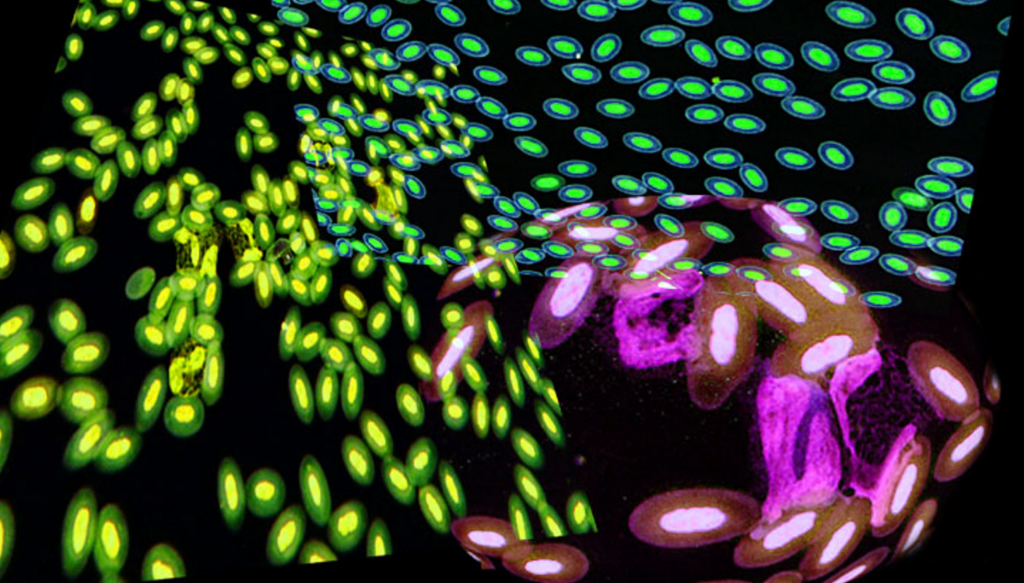
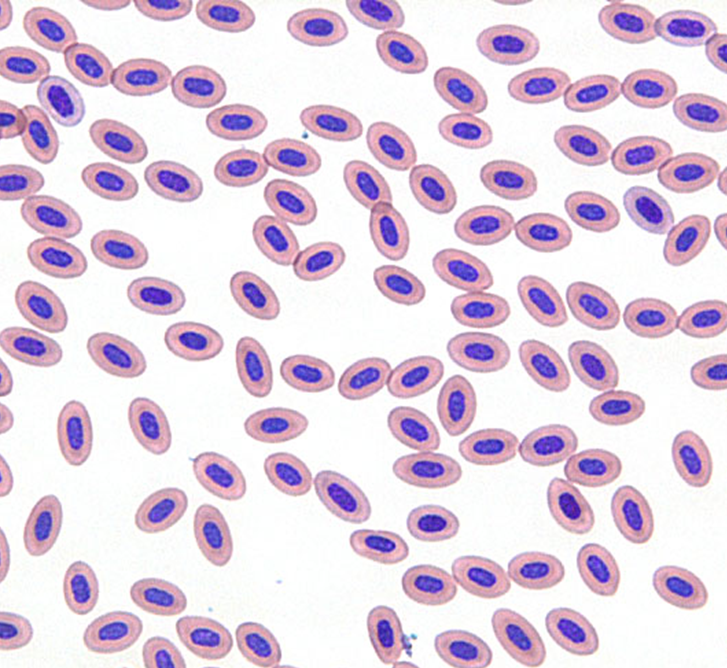
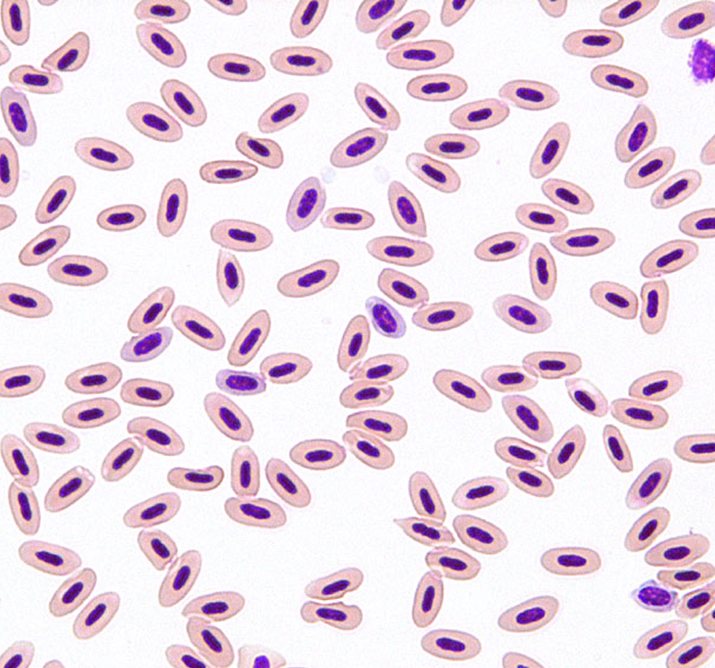
鳥類は高度に血管化されたが膨張しない肺と、ガス交換に関与しない無血管の気嚢を持ち、空気呼吸する他の脊椎動物とは非常に異なる呼吸器系を持っています〔Hill et al.2016〕。このような空気弁と一方向の空気の流れの存在は、内温性、特に飛行の高い有酸素運動要求を満たすためのメカニズムとして進化したと考えられてきました〔Farmer 2006〕。さらに心臓の大きさは飛行スタイルに関連すると推測されます〔Nespolo et al.2018〕。基本的には行動によって赤血球の形状ならびにサイズが異なる可能性があり、鳥種によって相違が見られます。以下のアヒルとセキセイインコの赤血球を観察すると、アヒルは丸く、セキセイインコは細長いです。
アヒルセキセイインコ
飛翔
持久飛行中の鳥は休息時の5~7倍の代謝率で活動します〔Engel et al.2006,Klaassen et al.2000,Lindström et al.1999〕。赤血球は高酸素流に継続的にさらされていると、酸化ダメージを受けやすく、細胞変性により血球寿命も短くなり、貧血が起こります〔Lutz et al.2013,Wagner et al.1987,Waugh et al.1992〕 。さらに、酸化ダメージは溶血を引き起こすことも知られており〔Lutz et al.2013,Sentürk et al.2001,Şentürk et al.2005〕。渡りの中継地に到着したオオソリハシシギ (Limosa lapponica taymyrensis) のヘマトクリット値とヘモグロビン含有量が低いことからわかるように、運動誘発性貧血は渡りの途中で発生する可能性があります〔Landys-Ciannelli et al.2002,Piersma et al.1996〕。
繁殖競争や渡りなどの活発な活動に従事することは、赤血球の大きさと伸長率に有意な影響を及ぼすことが分かりました〔Carl et al.2022〕。まず、渡りの途中で長距離を移動する鳥(および高緯度で繁殖する鳥)は、赤血球が小さく長いことがわかりました。赤血球が小さいほど、表面積と体積の比率が大きく、拡散距離が短いため、赤血球の体積が減少するにつれて、ヘモグロビンの酸素化と脱酸素化がより速くなります〔Holland et al,1966,Jones 1979〕。運動中に酸素を送達する能力は、有酸素能力の重要な要素で、これは、ガス交換領域の有効面積が大きく拡散距離が短いこと、心臓が大きいこと、飛翔筋の毛細血管密度が高いことなど、渡り鳥に見られるさまざまな生理学的適応と一致しています〔Lundgren et al.1988,McWilliams et al.2004)。赤血球が小さく長いと、筋肉の毛細血管系を通じた輸送も促進され〔Noguchi et al.2005〕、飛翔に必要なより効果的な持続的な有酸素運動が可能になります。
潜水種
潜水生活様式を持つ鳥種では、非潜水種よりも赤血球サイズが有意に大きいです〔Carl et al.2022〕。潜水行動はエネルギーコストが高いですが〔Butler 1991〕、空中での運動とは異なり、貯蔵酸素に依存しており、これは主に長時間の潜水では血液と筋肉に、短時間の潜水では呼吸器系に貯蔵されています〔Kooyman et al.1998〕。鳥類を含む潜水動物は血中ヘモグロビン濃度が高く〔Minias 2020〕、これが結合酸素を貯蔵する役割を果たしています。また、最も長い潜水を行うペンギンではヘモグロビンと酸素の親和性とボーア効果が増加しています〔Signore et al.2021〕。赤血球が大きいほど、より多くの酸素を結合するのに大きく貢献するため、酸素貯蔵容量にとって重要です。潜水哺乳類では、非潜水種よりも赤血球サイズが大きいことが示されており〔Wickham et al.1989〕、より大きな赤血球が潜水への適応として一般的に影響していることを裏付けています。
高山棲息種
高地(2000メートル以上)に生息する脊椎動物は、好気性代謝に問題となる低酸素状態にさらされます。鳥類は他の陸生脊椎動物よりも低酸素状態に耐性があります〔Lasiewski et al.1971,Faraci 1991〕。高地はより細長い赤血球と関連しており、高地で繁殖するアルプスのイワヒバリ(Prunella collaris)で細長い赤血球が報告されており〔Janiga et al. 2017,Haas et al.2020〕、楕円体はより大きな表面積を維持し、丸い細胞よりも効率的なガス交換を促進していました〔Hartman et al.1963〕。より細長い細胞は幅を広げることなく表面積を維持していますが、毛細血管床を通る流れを妨げる可能性があります。
繁殖・交配行動の影響
相対的に精巣サイズが大きい多くの鳥種は赤血球サイズが小さいことがわかり、社会的および遺伝的交配システムの両方に関係するため、オス同士の交尾のための競争が関与しているのかもしれません〔Soulsbury 2010,Soulsbury 2019〕 。実験的には個体にテストステロンを投与するとヘモグロビンレベルが上昇し、赤血球容積 (平均赤血球容積) が小さくなります 〔Puerta et al.1995〕。アンドロゲンレベルが高いほど赤血球生成率が上昇するため、丸い赤血球は循環血中の若い赤血球数の増加を反映している可能性があります〔Puerta et al. 1995〕 。いずれにせよ、酸素運搬能力はオスのパフォーマンスを強力に支えているため、性選択において重要な形質であること示唆しています。
Butler PJ.Exercise in birds.J Exp Biol160:233‐262.1991
Butler JP,Banzett RB,Fredberg JJ.Inspiratory valving in avian bronchi:aerodynamic considerations.Respir Physiol72:241-255.1988
Carl D Soulsbury CD,Dobson J,Deeming DC,Minias P.Energetic Lifestyle Drives Size and Shape of Avian Erythrocytes.Integrative and Comparative Biology62(1):p71-80.2022
Engel S,Biebach H,Visser GH.Metabolic costs of avian flight in relation to flight velocity:a study in rose coloured starlings (Sturnus roseus, Linnaeus). J Comp Physiol B Biochem Syst Environ Physiol176:415-427.2006
Faraci FM.Adaptations to hypoxia in birds: how to fly high.Annu Rev Physiol53:59-70.1991
Farmer CG.On the origin of avian air sacs.Respir Physiol Neurobiol154:89-106.2006
Glomski CA,Pica A.Erythrocytes of the Poikilotherms.London, UK:Foxwell & Davies UK Ltd.2006
Haas M,Janiga M.Variation in erythrocyte morphology in alpine accentors (Prunella collaris Scop.) from Tian Shan,Rila and the High Tatra mountains and effects of molting.Eur Zool J87:475-488.2020
Holland RAB,Forster RE.The effect of size of red cells on the kinetics of their oxygen uptake.J Gen Physiol49:727-742.1966
Hartman FA,Lessler MA..Erythrocyte measurements in birds.Auk80:467-473.1963
Hawkey CM,Bennett PM,Gascoyne SC,Hart MG,Kirkwood JK.Erythrocyte size, number and haemoglobin content in vertebrates.Br J Haematol77:392-397.1991
Hill RW,Wyse GA,Anderson M.Animal Physiology.4th ed.Sunderland (MA):Sinauer Associates Inc.Publishers.2016
Janiga M,Haas M.Alpine accentors as monitors of atmospheric long-range lead and mercury pollution in alpine environments.Environ Sci Pollut Res26:2445-2454.2019
Janiga M,Haas M,Kufelová M.Age,sex and seasonal variation in the shape and size of erythrocytes of the alpine accentor,Prunella collaris (Passeriformes: Prunellidae).Eur Zool J84:583–90.2017
Jones DA.The importance of surface area/volume ratio to the uptake of oxygen by red cells.J Gen Physiol74:643-646.1979
Klaassen M,Kvist A, Lindström Å.Flight costs and fuel composition of a bird migrating in a wind tunnel. Condor. 2000;102:444-51.2000
Kooyman GL,Ponganis PJ.The physiological basis of diving to depth:birds and mammals.Annu Rev Physiol60:19-32.1988
Kozłowski J,Czarnołęski M,François-Krassowska A,Maciak S, Pis T.Cell size is positively correlated between different tissues in passerine birds and amphibians, but not necessarily in mammals.Biol Lett 6:792-796.2010
Landys-Ciannelli MM,Jukema J,iersma T.Blood parameter changes during stopover in a long-distance migratory shorebird, the bar-tailed godwit Limosa lapponica taymyrensis.J Avian Biol33:451-455.2002
Lindström Å,Klaassen M,Kvist A. Variation in energy intake and basal metabolic rate of a bird migrating in a wind tunnel.Funct Ecol13:352-359.1999
Lutz HU,Bogdanova A.Mechanisms tagging senescent red blood cells for clearance in healthy humans.Front Physiol4:1-15.2013
Lundgren BO,Kiessling KH.Comparative aspects of fibre types,areas,and capillary supply in the pectoralis muscle of some passerine birds with differing migratory behaviour.J Comp Physiol B158:165–173.1998
Lasiewski RC,Calder WA Jr.A preliminary allometric analysis of respiratory variables in resting birds.Respir Physiol11:152-166.1971
Lahnsteiner F.Erythrocyte morphometry in teleost fish—species-specific, inter-individual and environmental-related differences.Acta Zool102:e64715.2020
McWilliams SR,Guglielmo C,Pierce B,Klaassen M.Flying, fasting, and feeding in birds during migration:a nutritional and physiological ecology perspective.J Avian Biol35:377-393.2004
Minias P.Ecology and evolution of blood oxygen-carrying capacity in birds.Am Nat195:788‐801.2020
Minias P,Kaczmarek K,Włodarczyk R,Janiszewski T.Hemoglobin concentrations in waders vary with their strategies of migration:a comparative analysis.Comp Biochem Physiol A: Mol Integr Physiol165:7-12.2013
Nespolo RF,González-Lagos C,Solano-Iguaran JJ,Elfwing M,Garitano-Zavala A,Mañosa S,Alonso JC,Altimiras J.Aerobic power and flight capacity in birds: a phylogenetic test of the heart-size hypothesis.J Exp Biol.221:jeb162693.2018
Noguchi H,Gompper G.Shape transitions of fluid vesicles and red blood cells in capillary flows. Proc Natl Acad Sci102:14159-164.2005
Piersma T,Everaarts JM,Jukema J. Build-up of red blood cells in refuelling bar-tailed Godwits in relation to individual migratory quality.Condor98:363–70.1996
Pis T.Resting metabolic rate and erythrocyte morphology in early development of thermoregulation in the precocial grey partridge (Perdix perdix).Comp Biochem Physiol A: Mol Integr Physiol151:211-218.2008
Ruiz G,Rosenmann M,Cortes A.Thermal acclimation and seasonal variations of erythrocyte size in the Andean mouse Phyllotis xanthopygus rupestris.Comp Biochem Physiol A:Mol Integr Physiol139:405-409.2004
Puerta M,Nava MP,Venero C,Veiga JP.Hematology and plasma chemistry of house sparrows (Passer domesticus) along the summer months and after testosterone treatment.Comp Biochem Physiol Part A Physiol110:303-307.1995
Signore AV,Tift MS,Hoffmann FG,Schmitt TL,Moriyama H,Storz JF.2021.Evolved increases in hemoglobin-oxygen affinity and the Bohr effect coincided with the aquatic specialization of penguins. Proc Natl Acad Sci118:e2023936118.2021
Sentürk ÜK,Gündüz F,Kuru O,Aktekin MR, Kipmen D,Yalçin Ö, Bor-Küçükatay M,Yeşilkaya A,Başkurt OK.Exercise-induced oxidative stress affects erythrocytes in sedentary rats but not exercise-trained rats.J Appl Physiol;91:1999-2004.2001
Şentürk ÜK,Gündüz F,Kuru O,Koçer G,Özkaya YG,Yeşilkaya A,Bor-Küçükatay M,Üyüklü M,Yalçin Ö,Başkurt OK.Exercise-induced oxidative stress leads hemolysis in sedentary but not trained humans.J Appl Physiol99:1434-1441.2005
Soulsbury CD.Genetic patterns of paternity and testes size in mammals.PLoS One.5:e9581.2010
Soulsbury CD.Income and capital breeding in males:energetic and physiological limitations on male mating strategies.J Exp Biol.222:jeb184895.2019
Starostová Z,Konarzewski M,Kozłowski J,Kratochvíl L.Ontogeny of metabolic rate and red blood cell size in eyelid geckos: species follow different paths.PLoS One8:e64715.2013
Wagner G,Chiu D,Qju J,Heath R,Lubin B.Spectrin oxidation correlates with membrane vesiculation in stored RBCs.Blood69:1777-1781.1987
Waugh RE,Narla M,Jackson CW,Mueller TJ,Suzuki T,Dale GL.Rheologic properties of senescent erythrocytes:loss of surface area and volume with red blood cell age.Blood79:1351-1358.1992
Wickham LL,Elsner R,White FC,Cornell LH.Blood viscosity in phocid seals: possible adaptations to diving.J Comp Physiol B159:153-158.1989
Yap KN,Tsai OHI,Williams TD.Haematological traits co-vary with migratory status,altitude and energy expenditure:a phylogenetic,comparative analysis.Sci Rep9:6351.2019