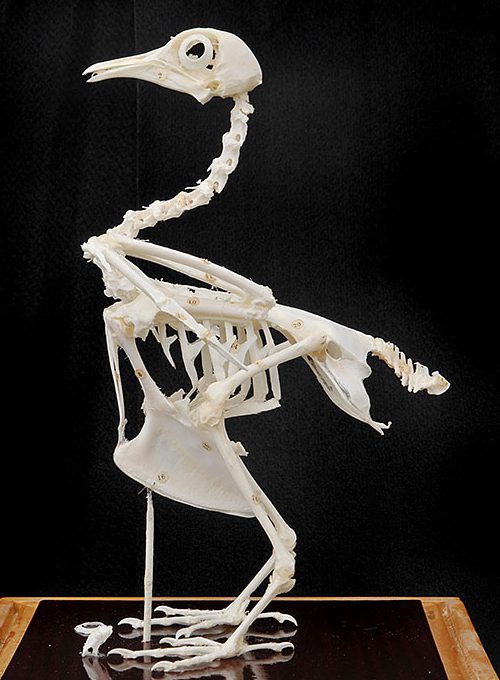
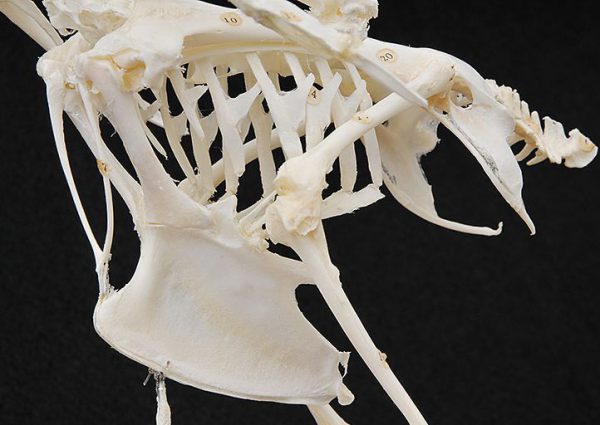
肋骨は胸椎と胸骨とともに強固な胸郭を形成しており、呼吸のために軽度に拡張すること もできます〔鮫島ら 1984〕。肋骨の脊椎と胸骨との間に蝶番があり、肋間筋の収縮を可能にし、鳥特有の呼吸様式である鞴呼吸を可能とします。また、鳥類のそれぞれの肋骨には鉤状突起があり、これらは鉤状に伸びた突起が、後ろの肋骨と重なることで胸郭のを支えて強化し〔Nakatsukasa 2004,Kardong 1988〕、呼吸の動きにも関与します〔Welty 1979〕。鉤状突起はカモ目のサケビドリ科などを除く、多くの鳥種に存在しますが〔Fowler et al.2001〕、突起の形状は鳥種によって異なり、飛翔しない鳥では小さく、潜水種では長いです〔Tickle et al.2007, Tickle et al.2009〕。
腰帯
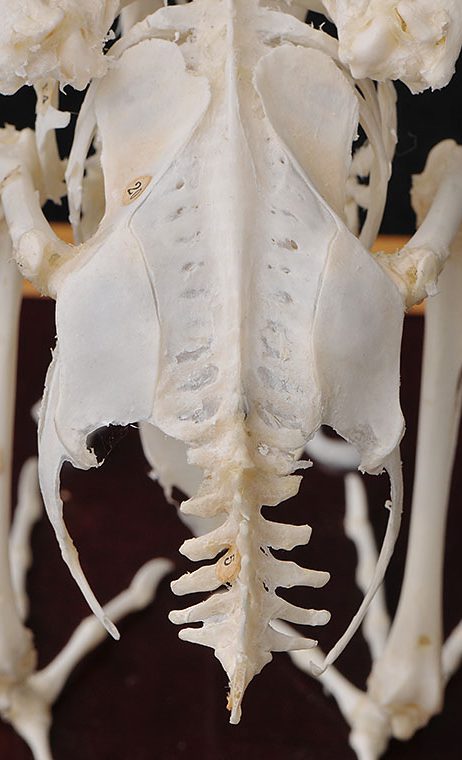
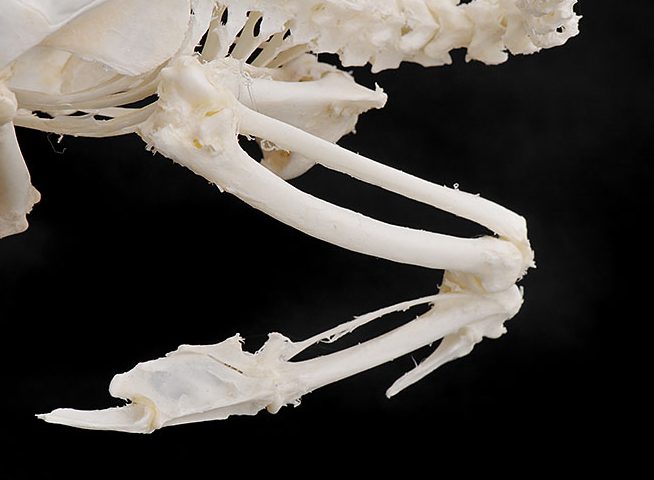
腰椎は個々の椎骨として存在せず、数個の胸椎と全腰椎、全仙椎、数個の尾椎が癒合して複合仙骨を形成します。さらに寛骨が両側から加わって癒合したものを腰仙骨と呼ぶこともあり、寛骨は腸骨、恥骨、坐骨が 一体化しています。恥骨は座骨と平行に後方に向かってお り、ダチョウを除き,左右の恥骨が結合せずに離れているのが特徴です〔Campbell et al.1985〕。鳥類においては左右の恥骨間距離が産卵のため、メスでオスより大きいことが知られており、恥骨間距離による雌雄鑑別が、特にオウムで行われています〔Stromberg 1977〕。尾椎の先端にはいくつかの尾椎が癒合した尾端骨が存在し、尾羽を支えて、可動するにも役立っています。
Marshall AJ.Biology and Comparative Physiology of Birds:Volume I.Victoria.Academic Press.Australia:p245-251.1960
Nakatsukasa M.Acquisition of bipedalism: the Miocene hominoid record and modern analogues for bipedal protohominids.Journal of Anatomy204(5):385‐402.2004
Stromberg L.(Compiled) .Sexing all fowls,baby chicks,game birds,cagebirds.Stromberg Publishing Company.Pine River.Minnesota.1975
Smith ND.Body mass and foraging ecology predict evolutionary patterns of skeletal pneumaticity in the diverse waterbird clade. Evolution66(4):1059-1078.2011
Tickle PG,Codd JR.Ontogenetic development of the uncinate processes in the domestic turkey. (Meleagris gallopavo).Poultry Sci88:179-184.2009
Tickle PG,Ennos AR,Lennox LE,Perry SF,Codd JR.Functional significance of the uncinate processes in birds.J.Exp.Biol210:3955–3961.2007
Wedel MJ.Vertebral pneumaticity,air sacs,and the physiology of sauropod dinosaurs (PDF).Paleobiology29(2):243-255.2003
Welty JC.The Life of Birds 2nd edtn.Saunders.Philadelphia.1979